Для изучения процессов, протекающих в организме, нужно знать, что происходит на клеточном уровне. А там важнейшую роль играют белковые соединения. Необходимо изучить не только их функции, но и процесс создания. Поэтому важно объяснить кратко и понятно. 9 класс для этого подходит самым лучшим образом. Именно на этом этапе учащиеся владеют достаточным количеством знаний для понимания данной темы.
Белки - что это такое и для чего они нужны
Эти высокомолекулярные соединения играют огромную роль в жизни любого организма. Белки являются полимерами, то есть состоят из множества похожих «кусочков». Их количество может варьироваться от нескольких сотен до тысяч.
В клетке белки выполняют множество функций. Велика их роль и на более высоких уровнях организации: ткани и органы во многом зависят от правильной работы различных белков.
Например, все гормоны имеют белковое происхождение. А ведь именно эти вещества контролируют все процессы в организме.
Гемоглобин - тоже белок, он состоит из четырех цепей, которые в центре соединены атомом железа. Такая структура обеспечивает возможность переносить кислород эритроцитами.
Напомним, что все мембраны имеют в своем составе белки. Они необходимы для переноса веществ сквозь оболочку клеток.
Существует еще множество функций белковых молекул, которые они выполняют четко и беспрекословно. Эти удивительные соединения очень разнообразны не только по своим ролям в клетке, но и по строению.
Где происходит синтез
Рибосома является органеллой, в которой проходит основная часть процесса, называемого "биосинтез белка". 9 класс в разных школах отличается по программе изучения биологии, но многие учителя дают материал по органеллам заблаговременно, до изучения трансляции.
Поэтому учащимся будет нетрудно вспомнить пройденный материал и закрепить его. Следует знать, что на одной органелле одновременно может создаваться только одна полипептидная цепь. Этого мало, чтобы удовлетворить все потребности клетки. Поэтому рибосом очень много, и чаще всего они объединяются с эндоплазматической сетью.

Такая ЭПС называется шероховатой. Выгода такого «сотрудничества» очевидна: белок сразу после синтеза попадает в транспортный канал и может без задержек отправляться в место назначения.
Но если принимать во внимание самое начало, а именно считывание информации с ДНК, то можно сказать, что биосинтез белка в живой клетке начинается еще в ядре. Именно там синтезируется которая содержит генетический код.
Необходимые материалы - аминокислоты, место синтеза - рибосома
Кажется, что сложно объяснить, как протекает биосинтез белка, кратко и понятно, схема процесса и многочисленные рисунки просто необходимы. Они помогут донести всю информацию, а также учащимся удастся легче ее запомнить.
Прежде всего, для синтеза необходим «строительный материал» - аминокислоты. Некоторые из них вырабатываются организмом. Другие же можно получить только с пищей, они называются незаменимыми.

Общее число аминокислот - двадцать, но за счет огромного числа вариантов, в которых можно их располагать в длинной цепочке, молекулы белков очень разнообразны. Эти кислоты похожи между собой по структуре, но отличаются радикалами.
Именно свойства этих частей каждой аминокислоты определяют, в какую структуру «свернется» получившаяся цепочка, будет ли она образовывать четвертичную структуру с другими цепями, и какими свойствами будет обладать получившаяся макромолекула.

Процесс биосинтеза белка не может протекать просто в цитоплазме, для него нужна рибосома. состоит из двух субъединиц - большой и малой. В состоянии покоя они разобщены, но как только начинается синтез, они сразу соединяются и начинают работать.
Такие разные и важные рибонуклеиновые кислоты
Для того чтобы принести аминокислоту к рибосоме, нужна специальная РНК, называемая транспортной. Для сокращения ее обозначают т-РНК. Эта одноцепочечная молекула в виде клеверного листа способна прицепить одну аминокислоту к своему свободному концу и переправить ее к месту синтеза белка.
Еще одна РНК, участвующая в синтезе белка, называется матричной (информационной). Она несет в себе не менее важный компонент синтеза - код, в котором четко прописано, когда какую аминокислоту цеплять к образующейся цепочке белка.
Эта молекула имеет одноцепочечное строение, состоит из нуклеотидов, так же как и ДНК. Существуют некоторые отличия в первичной структуре этих нуклеиновых кислот, о которых вы можете прочитать в сравнительной статье о РНК и ДНК.
Информацию о составе белка м-РНК получает от главного хранителя генетического кода - ДНК. Процесс чтения и синтеза м-РНК называется транскрипцией.
Он происходит в ядре, откуда получившаяся м-РНК отправляется к рибосоме. Сама же ДНК из ядра не выходит, ее задача - только сохранить генетический код и передать его дочерней клетке во время деления.
Сводная таблица главных участников трансляции
Для того чтобы описать биосинтез белка кратко и понятно, таблица просто необходима. В нее мы запишем все компоненты и их роль в этом процессе, который называется трансляцией.
Сам же процесс создания белковой цепочки делится на три этапа. Давайте рассмотрим каждый из них более подробно. После этого вы сможете легко объяснить всем желающим биосинтез белка кратко и понятно.
Инициация - начало процесса
Это начальная стадия трансляции, в которой малая субъединица рибосомы соединяется с самой первой т-РНК. Эта рибонуклеиновая кислота несет на себе аминокислоту - метионин. Трансляция всегда начинается именно с этой аминокислоты, так как стартовым кодоном является АУГ, который и кодирует этот первый мономер в белковой цепи.
Для того чтобы рибосома узнала стартовый кодон и не начала синтез с середины гена, где последовательность АУГ тоже может оказаться, вокруг начального кодона располагается специальная последовательность нуклеотидов. Именно по ним рибосома узнает то место, на которое должна сесть ее малая субъединица.
После образования комплекса с м-РНК, стадия инициации заканчивается. И начинается основной этап трансляции.
Элонгация - середина синтеза
На этом этапе происходит постепенное наращивание белковой цепочки. Продолжительность элонгации зависит от количества аминокислот в белке.

Первым делом к малой субъединице рибосомы присоединяется большая. И начальная т-РНК оказывается в ней целиком. Снаружи остается только метионин. Далее в большую субъединицу заходит вторая т-РНК, несущая другую аминокислоту.
Если второй кодон на м-РНК совпадает с антикодоном на верхушке «клеверного листа», вторая аминокислота присоединяется к первой с помощью пептидной связи.
После этого рибосома передвигается по м-РНК ровно на три нуклеотида (один кодон), первая т-РНК отсоединяет от себя метионин и отделяется от комплекса. На ее месте оказывается вторая т-РНК, на конце которой висит уже две аминокислоты.
Затем в большую субъединицу входит третья т-РНК и процесс повторяется. Он будет происходить до тех пор, пока рибосома не наткнется на кодон в м-РНК, который сигнализирует об окончании трансляции.
Терминация
Этот этап является последним, некоторым он может показаться весьма жестоким. Все молекулы и органеллы, которые так слаженно работали над созданием полипептидной цепочки, останавливаются, как только рибосома наезжает на терминальный кодон.
Он не кодирует ни одну аминокислоту, поэтому какая бы т-РНК ни зашла в большую субъединицу, все они будут отвергнуты из-за несоответствия. Тут в дело вступают факторы терминации, которые отделяют готовый белок от рибосомы.

Сама органелла может либо распасться на две субъединицы, либо продолжить свой путь по м-РНК в поисках нового стартового кодона. На одной м-РНК могут находиться сразу несколько рибосом. Каждая из них - на свой стадии трансляции.Только что созданный белок снабжается маркерами, с помощью которых всем будет понятно его место назначения. И по ЭПС он будет отправлен туда, где необходим.
Чтобы понять роль биосинтеза белка, необходимо изучить, какие функции он может выполнять. Это зависит от последовательности аминокислот в цепочке. Именно их свойства определяют вторичную, третичную, а иногда и четвертичную (если она существует) и его роль в клетке. Более подробно о функциях белковых молекул можно прочитать в статье по этой теме.
Как узнать больше о трансляции
В этой статье описан биосинтез белка в живой клетке. Конечно, если изучать предмет глубже, на объяснение процесса во всех подробностях уйдет немало страниц. Но вышеизложенного материала должно хватить для общего представления.Очень полезным для понимания могут оказаться видеоматериалы, в которых ученые смоделировали все этапы трансляции. Некоторые из них переведены на русский язык и могут послужить отличным пособием для учащихся или просто познавательным видео.

Для того чтобы разбираться в теме лучше, следует прочитать и другие статьи на близкие темы. Например, про или про функции белков.
В каждой области науки есть своя «синяя птица»; кибернетики мечтают о «думающих» машинах, физики - об управляемых термоядерных реакциях, химики - о синтезе «живого вещества» - белка. Синтез белка долгие годы был темой фантастических романов, символом грядущего могущества химии. Это объясняется и той огромной ролью, какая принадлежит белку в мире живого, и теми трудностями, которые неизбежно вставали перед каждым смельчаком, отважившимся «сложить» из отдельных аминокислот замысловатую мозаику белка. И даже еще не самого белка, а только пептидов.
Разница между белками и пептидами не только терминологическая, хотя молекулярные цепи и тех и других состоят из аминокислотных остатков. На каком-то этапе количество переходит в качество: пептидная цепь - первичная структура - обретает способность сворачиваться в спирали и клубки, образуя вторичную и третичную структуры, характерные уже для живой материи. И тогда пептид становится белком. Четкой границы здесь не существует - на полимерной цепи нельзя поставить демаркационный знак: досель - пептид, отсель - белок. Но известно, например, что адранокортикотропный гормон, состоящий из 39 остатков аминокислот,- это полипептид, а гормон инсулин, состоящий из 51 остатка в виде двух цепей,- это уже белок. Простейший, но все же белок.
Способ соединения аминокислот в пептиды был открыт в начале прошлого века немецким химиком Эмилем Фишером. Но еще долго после этого химики не могли всерьез помышлять не только о синтезе белка или 39-членных пептидов, но даже значительно более коротких цепей.
Процесс синтеза белка
Для того, чтобы соединить между собой две аминокислоты, надо преодолеть немало трудностей. Каждая аминокислота, подобно двуликому Янусу, имеет два химических лица: карбоксильную кислотную группу на одном конце и аминную основную группу - на другом. Если от карбоксила одной аминокислоты отнять группу ОН, а от аминной группы другой - атом , то образовавшиеся при этом два аминокислотных остатка могут соединиться друг с другом пептидной связью, и в результате возникнет простейший из пептидов - дипептид. И отщепится молекула воды. Повторяя эту операцию, можно наращивать длину пептида.
Однако эта, казалось бы, на первый взгляд несложная операция практически трудноосуществима: аминокислоты очень неохотно соединяются друг с другом. Приходится их активировать, химически, и «подогревать» один из концов цепи (чаще всего карбоксильный), и вести реакцию, строго соблюдая необходимые условия. Но это еще не все: вторая сложность состоит в том, что соединяться друг с другом могут не только остатки разных аминокислот, но и две молекулы одной кислоты. При этом строение синтезируемого пептида будет уже отличаться от желаемого. Больше того, каждая аминокислота может иметь не две, а несколько «ахиллесовых пят» - боковых химически активных групп, способных присоединять аминокислотные остатки.
Чтобы не дать реакции свернуть с заданного пути, необходимо закамуфлировать эти ложные мишени - «запечатать» на время осуществляемой реакции все реакционноспособные группы аминокислоты, кроме одной, присоединив к ним так называемые защитные группировки. Если этого не сделать, то цель будет расти не только с обоих концов, но и вбок, и аминокислоты уже не удастся соединить в заданной последовательности. А ведь именно в этом и заключается смысл всякого направленного синтеза.
Но, избавляясь таким образом от одной неприятности, химики столкнулись с другой: защитные группировки после окончания синтеза нужно удалить. Во времена Фишера в качестве «защиты» применялись группировки, которые отщеплялись гидролизом. Однако реакция гидролиза обычно оказывалась слишком сильным «потрясением» для полученного пептида: с трудом построенная его «конструкция» разваливалась как только с нее снимали «строительные леса» - защитные группировки. Лишь в 1932 году ученик Фишера М. Бергманн нашел выход из этого положения: он предложил защищать аминогруппу аминокислоты карбобензоксигруппой, которую можно было удалить без повреждения пептидной цепи.
Синтез белка из аминокислот
В течение последующих лет был предложен ряд так называемых мягких методов «сшивки» аминокислот друг с другом. Однако все они фактически были лишь вариациями на тему метода Фишера. Вариациями, в которых иногда даже трудно было уловить исходную мелодию. Но сам принцип оставался все тем же. И все теми же оставались трудности, связанные с защитой уязвимых групп. За преодоление этих трудностей приходилось расплачиваться увеличением числа стадий реакции: один элементарный акт - соединение двух аминокислот - распадался на четыре этапа. А каждая лишняя стадия - это неизбежные потери.
Если даже предположить, что каждая стадия идет с полезным выходом в 80% (а это хороший выход), то через четыре этапа эти 80% «растают» до 40%. И это при синтезе только дипептида! А если аминокислот будет 8? А если 51, как в инсулине? Прибавьте к этому сложности, связанные с существованием двух оптических «зеркальных» форм молекул аминокислот, из которых в реакции нужна только одна, приплюсуйте проблемы отделения образующихся пептидов от побочных продуктов, особенно в тех случаях, когда они одинаково растворимы. Что же получится в сумме: Дорога в никуда?
И все же эти трудности не останавливали химиков. Погоня за «синей птицей» продолжалась. В 1954 году были синтезированы первые биологически активные гормоны-полипептиды - вазопрессин и окситоцин. В них было по восемь аминокислот. В 1963 году был синтезирован 39-членный полипептид АКТГ - адренокортикотропный гормон. Наконец, химики США, Германии и Китая синтезировали первый белок - гормон инсулин.
Как же так, скажет читатель, трудная дорога, оказывается, привела не в никуда и не куда-нибудь, а к осуществлению мечты многих поколений химиков! Это же эпохальное событие! Верно, это - эпохальное событие. Но давайте оценим его трезво, отрешившись от сенсационности, восклицательных знаков и чрезмерных эмоций.
Никто не спорит: синтез инсулина - огромная победа химиков. Это колоссальный, титанический труд, достойный всякого восхищения. Но вместе с тем эго, по существу, и потолок старой химии полипептидов. Это победа на грани поражения.
Синтез белков и инсулин
В инсулине 51 аминокислота. Чтобы соединить их в нужной последовательности, химикам потребовалось провести 223 реакции. Когда спустя три года после начала первой из них была закончена последняя, выход продукта составлял меньше одной сотой процента. Три года, 223 стадии, сотая доля процента - согласитесь, победа носит чисто символический характер. Говорить о практическом применении этого метода очень трудно: слишком велики связанные с его реализацией расходы. А ведь в конечном счете речь идет о синтезе не драгоценных реликвий славы органической химии, а о выпуске жизненно важного лекарственного препарата, который необходим тысячам людей во всем мире. Так классический метод синтеза полипептидов исчерпал себя на первом же, самом простом белке. Значит, «синяя птица» вновь ускользнула из рук химиков?
Новый метод синтеза белка
Примерно за полтора года до того, как мир узнал о синтезе инсулина, в печати промелькнуло еще одно сообщение, которое вначале не привлекло особого внимания: американский ученый Р. Мэрифилд предложил новый метод синтеза пептидов. Поскольку сам автор поначалу не дал методу должной оценки, и в нем было много недоработок, выглядел он в первом приближении даже хуже существовавших. Однако уже в начале 1964 года, когда Мэрифилду удалось с помощью своего метода осуществить полный синтез 9-членного гормона с полезным выходом в 70%, ученые изумились: 70% после всех этапов - это 9% полезного выхода на каждой стадии синтеза.
Основная идея нового метода заключается в том, что растущие цепочки пептидов, которые раньше были брошены на произвол хаотического движения в растворе, теперь привязывались одним концом к твердому носителю - их как бы заставляли стать на якорь в растворе. Мэрифилд брал твердую смолу и к ее активным группам «привязывал» за карбонильный конец первую из собираемых в пептид аминокислоту. Реакции шли внутри отдельных частичек смолы. В «лабиринтах» ее молекул сначала появлялись первые короткие ростки будущего пептида. Затем в сосуд вводили вторую аминокислоту, ее молекулы сшивались своими карбонильными концами со свободными аминными концами «привязанной» аминокислоты, и в частицах вырастал еще один «этаж» будущего «здания» пептида. Так, этап за этапом, постепенно наращивался весь пептидный полимер.
Новый метод имел несомненные преимущества: прежде всего в нем была решена проблема отделения ненужных продуктов после присоединения каждой очередной аминокислоты - эти продукты легко смывались, а пептид оставался пришитым к гранулам смолы. Одновременно исключалась проблема растворимости растущих пептидов - один из главных бичей старого метода; раньше они нередко выпадали в осадок, практически переставая участвовать в процессе роста. Пептиды, «снимаемые» после окончания синтеза с твердой подложки, получались почти все одинакового размера и строения, во всяком случае, разброс в структуре был меньше, чем при классическом методе. И соответственно больше полезный выход. Благодаря этому методу синтез пептидов - кропотливый, трудоемкий синтез - легко поддается автоматизации.
Мэрифилд соорудил несложный автомат, который сам по заданной программе проделывал все положенные операции - подачу реагентов, смешивание, слив, промывку, отмер дозы, добавление новой порции и так далее. Если по старому методу на присоединение одной аминокислоты приходилось травить 2-3 дня, то Мэрифилд на своем автомате соединял за день 5 аминокислот. Разница - в 15 раз.
В чем состоят трудности синтеза белков
Метод Мэрифилда, названный твердофазным, или гетерогенным, сразу же был принят на вооружение химиками всего мира. Однако уже через короткое время стало ясно: новый метод вместе с крупными достоинствами имеет и ряд серьезных недостатков.
По мере роста пептидных цепей может случиться так, что в какой-то из них окажется пропущенным, скажем, третий «этаж» - третья по счету аминокислота: ее молекула не дойдет до места соединения, застряв где-нибудь по дороге в структурных «дебрях» твердого полимера. И тогда, даже если все остальные аминокислоты, начиная с четвертой, выстроятся в должном порядке, это уже не спасет положения. Полученный полипептид по своему составу, а следовательно, и по своим свойствам не будет иметь ничего общего с получаемым веществом. Произойдет то же самое, что и при наборе телефонного номера; стоит пропустить одну цифру - и нам уже не поможет тот факт, что все остальные мы набрали правильно. Отделить же такие ложные цепи от «настоящих» практически невозможно, и препарат оказывается засоренным примесями. Кроме того, оказывается, что синтез нельзя вести на какой угодно смоле - ее нужно тщательно подбирать, так как свойства растущего пептида зависят в какой-то мере от свойств смолы. Поэтому ко всем этапам синтеза белка необходимо подходить максимально тщательно.
Синтез белка ДНК, видео
И под конец, предлагаем вашему вниманию образовательное видео о том, как происходит синтез белка в молекулах ДНК.
Биосинтез белков является важнейшим процессом анаболизма. Все признаки, свойства и функции клеток и организмов определяются в конечном итоге белками. Белки недолговечны, время их существования ограничено. В каждой клетке постоянно синтезируются тысячи различных белковых молекул. В начале 50-х гг. ХХ в. Ф. Крик сформулировал центральную догму молекулярной биологии: ДНК → РНК → белок. Согласно этой догме способность клетки синтезировать определенные белки закреплена наследственно, информация о последовательности аминокислот в белковой молекуле закодирована в виде последовательности нуклеотидов ДНК. Участок ДНК, несущий информацию о первичной структуре конкретного белка, называетсягеном . Гены не только хранят информацию о последовательности аминокислот в полипептидной цепочке, но и кодируют некоторые виды РНК: рРНК, входящие в состав рибосом, и тРНК, отвечающие за транспорт аминокислот. В процессе биосинтеза белка выделяют два основных этапа: транскрипция - синтез РНК на матрице ДНК (гена) - и трансляция - синтез полипептидной цепи.
Генетический код и его свойства
Генетический код - система записи информации о последовательности аминокислот в полипептиде последовательностью нуклеотидов ДНК или РНК. В настоящее время эта система записи считается расшифрованной.
Свойства генетического кода:
- триплетность: каждая аминокислота кодируется сочетанием из трех нуклеотидов (триплетом, кодоном);
- однозначность (специфичность): триплет соответствует только одной аминокислоте;
- вырожденность (избыточность): аминокислоты могут кодироваться несколькими (до шести) кодонами;
- универсальность: система кодирования аминокислот одинакова у всех организмов Земли;
- неперекрываемость: последовательность нуклеотидов имеет рамку считывания по 3 нуклеотида, один и тот же нуклеотид не может быть в составе двух триплетов;
- из 64 кодовых триплетов 61 - кодирующие, кодируют аминокислоты, а 3 - бессмысленные (в РНК - УАА, УГА, УАГ), не кодируют аминокислоты. Они называются кодонами-терминаторами , поскольку блокируют синтез полипептида во время трансляции. Кроме того, есть кодон-инициатор (в РНК - АУГ), с которого трансляция начинается.
| Первое основание | Второе основание | Третье основание |
|||
|---|---|---|---|---|---|
| У(А) | Ц(Г) | А(Т) | Г(Ц) | ||
| У(А) | Фен Фен Лей Лей | Сер Сер Сер Сер | Тир Тир - - | Цис Цис - Три | У(А) Ц(Г) А(Т) Г(Ц) |
| Ц(Г) | Лей Лей Лей Лей | Про Про Про Про | Гис Гис Глн Глн | Арг Арг Арг Арг | У(А) Ц(Г) А(Т) Г(Ц) |
| А(Т) | Иле Иле Иле Мет | Тре Тре Тре Тре | Асн Асн Лиз Лиз | Сер Сер Арг Арг | У(А) Ц(Г) А(Т) Г(Ц) |
| Г(Ц) | Вал Вал Вал Вал | Ала Ала Ала Ала | Асп Асп Глу Глу | Гли Гли Гли Гли | У(А) Ц(Г) А(Т) Г(Ц) |
* Первый нуклеотид в триплете - один из четырех левого вертикального ряда, второй - один из верхнего горизонтального ряда, третий - из правого вертикального.
Реакции матричного синтеза
Это особая категория химических реакций, происходящих в клетках живых организмов. Во время этих реакций происходит синтез полимерных молекул по плану, заложенному в структуре других полимерных молекул-матриц. На одной матрице может быть синтезировано неограниченное количество молекул-копий. К этой категории реакций относятся репликация, транскрипция, трансляция и обратная транскрипция.
Ген - участок молекулы ДНК, кодирующий первичную последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекулах транспортных и рибосомных РНК. ДНК одной хромосомы может содержать несколько тысяч генов, которые располагаются в линейном порядке. Место гена в определенном участке хромосомы называется локусом . Особенностями строения гена эукариот являются: 1) наличие достаточно большого количества регуляторных блоков, 2) мозаичность (чередование кодирующих участков с некодирующими).
Экзоны (Э) - участки гена, несущие информацию о строении полипептида.
Интроны (И) - участки гена, не несущие информацию о строении полипептида. Число экзонов и интронов различных генов разное; экзоны чередуются с интронами, общая длина последних может превышать длину экзонов в два и более раз. Перед первым экзоном и после последнего экзона находятся нуклеотидные последовательности, называемые соответственно лидерной (ЛП) и трейлерной последовательностью (ТП). Лидерная и трейлерная последовательности, экзоны и интроны образуют единицу транскрипции.
Промотор (П) - участок гена, к которому присоединяется фермент РНК-полимераза, представляет собой особое сочетание нуклеотидов. Перед единицей транскрипции, после нее, иногда в интронах находятся регуляторные элементы (РЭ), к которым относятся энхансеры и сайленсеры . Энхансеры ускоряют транскрипцию, сайленсеры тормозят ее.
Транскрипция - синтез РНК на матрице ДНК. Осуществляется ферментом РНК-полимеразой.
РНК-полимераза может присоединиться только к промотору, который находится на 3′-конце матричной цепи ДНК, и двигаться только от 3′- к 5′-концу этой матричной цепи ДНК. Синтез РНК происходит на одной из двух цепочек ДНК в соответствии с принципами комплементарности и антипараллельности. Строительным материалом и источником энергии для транскрипции являются рибонуклеозидтрифосфаты (АТФ, УТФ, ГТФ, ЦТФ).
В результате транскрипции образуется «незрелая» иРНК (про-иРНК), которая проходит стадию созревания или процессинга. Процессинг включает в себя: 1) КЭПирование 5′-конца, 2) полиаденилирование 3′-конца (присоединение нескольких десятков адениловых нуклеотидов), 3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиадениловый «хвост».
Транслируемая область начинается кодоном-инициатором, заканчивается кодонами-терминаторами. НТО содержат информацию, определяющую поведение РНК в клетке: срок «жизни», активность, локализацию.
Транскрипция и процессинг происходят в клеточном ядре. Зрелая иРНК приобретает определенную пространственную конформацию, окружается белками и в таком виде через ядерные поры транспортируется к рибосомам; иРНК эукариот, как правило, моноцистронны (кодируют только одну полипептидную цепь).
Трансляция
Трансляция - синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию, - рибосомы. У эукариот рибосомы находятся в некоторых органоидах - митохондриях и пластидах (70S-рибосомы), в свободном виде в цитоплазме (80S-рибосомы) и на мембранах эндоплазматической сети (80S-рибосомы). Таким образом, синтез белковых молекул может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для собственных нужд клетки; белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. В рибосоме выделяют малую и большую субъединицы. Малая субъединица рибосомы отвечает за генетические, декодирующие функции; большая - за биохимические, ферментативные.
В малой субъединице рибосомы расположен функциональный центр (ФЦР) с двумя участками - пептидильным (Р-участок) иаминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три - в пептидильном и три - в аминоацильном участках.
Для транспорта аминокислот к рибосомам используются транспортные РНК, тРНК. Длина тРНК от 75 до 95 нуклеотидных остатков. Они имеют третичную структуру, по форме напоминающую лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3′-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ). Таким образом, у каждой аминокислоты есть свои тРНК и свои ферменты, присоединяющие аминокислоту к тРНК.
Двадцать видов аминокислот кодируются 61 кодоном, теоретически может быть 61 вид тРНК с соответствующими антикодонами. Но кодируемых аминокислот всего 20 видов, значит, у одной аминокислоты может быть несколько тРНК. Установлено существование нескольких тРНК, способных связываться с одним и тем же кодоном (последний нуклеотид в антикодоне тРНК не всегда важен), поэтому в клетке обнаружено всего около 40 различных тРНК.
Синтез белка начинается с того момента, когда к 5′-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК (транспортирующая аминокислоту метионин). Следует отметить, что любая полипептидная цепь на N-конце сначала имеет метионин, который в дальнейшем чаще всего отщепляется. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислот.
Затем происходит присоединение большой субъединицы рибосомы, и в А-участок поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
Как только образовалась пептидная связь, метиониновая тРНК отсоединяется от метионина, а рибосома передвигается на следующий кодовый триплет иРНК, который оказывается в А-участке рибосомы, а метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами.
Трансляция идет до тех пор, пока в А-участок не попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения. Полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Скорость передвижения рибосомы по иРНК - 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут. Первым белком, синтезированным искусственно, был инсулин, состоящий из 51 аминокислотного остатка. Потребовалось провести 5000 операций, в работе в течение трех лет принимали участие 10 человек.
В трансляции можно выделить три стадии: а) инициации (образование иницаторного комплекса), б) элонгации (непосредственно «конвейер», соединение аминокислот друг с другом), в) терминации (образование терминирующего комплекса).
Транскрипция и трансляция у прокариот
«Механизмы» сборки полинуклеотидных и полипептидных цепочек у прокариот и эукариот не различаются. Но в связи с тем, что гены прокариот не имеют экзонов и интронов (исключение - гены архебактерий), располагаются группами, и на эту группу генов приходится один промотор, появляются следующие особенности транскрипции и трансляции у прокариот.
- В результате транскрипции образуется полицистронная иРНК, кодирующая несколько белков, совместно обеспечивающих определенную группу реакций.
- иРНК имеет несколько центров инициации трансляции, терминации трансляции и НТО.
- Не происходят КЭПирование, полиаденилирование и сплайсинг иРНК.
- Трансляция начинается еще до завершения транскрипции; эти процессы не разделены во времени и пространстве, как это имеет место у эукариот.
Можно добавить, что срок «жизни» прокариотических иРНК - несколько минут (у эукариот - часы и даже сутки).
Если видео не работает. Дополнительный
Одно из наиболее важных и характерных свойств живой клетки. Первичная структура белка, как уже отмечалось, предопределяется генетическим кодом, заложенным в молекуле ДНК, причем различные ее участки кодируют синтез разных белков. Следовательно, одна молекула ДНК хранит информацию о структуре многих белков.
Свойства белка зависят от последовательности расположения аминокислот в полипептидной цепи. В свою очередь чередование аминокислот определяется последовательностью нуклеотидов в ДНК. В иРНК каждой аминокислоте соответствует определенный триплет - группа, состоящая из трех нуклеотидов, называемая кодоном.
Биосинтез белка начинается в ядре со списывания информации о структуре белковой молекулы с ДНК на иРНК по принципу комплементарности. Данный процесс протекает как реакция матричного синтеза и называется транскрипцией (рис. 7.1).
Рис. 7.1. Процесс транскрипции
В результате транскрипции образуется «незрелая» иРНК (пре-иРНК), которая проходит стадию созревания или процессинга .
Процессинг включает в себя:
1) КЭПирование 5"-конца;
2) полиаденилирование 3"-конца (присоединение нескольких десятков адениловых нуклеотидов);
3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиА «хвост». Возможен альтернативный сплайсинг , при котором вместе с интронами вырезаются и экзоны. При этом с одного гена могут образовываться разные белки. Таким образом, утверждение – «Один ген – один полипептид» – неверно (рис. 7.2, 7.3, 7.4)

Рис. 7.2. Сплайсинг

Рис. 7.3. Альтернативный сплайсинг (варианты)

Рис. 7.4. Образование разных молекул белка при вариантах альтернативного сплайсинга
Образующаяся при этом иРНК поступает в цитоплазму, где на нее нанизываются рибосомы. Одновременно в цитоплазме с помощью ферментов активизируется транспортная РНК тРНК.
Молекула тРНК напоминает по структуре лист клевера, на вершине которого находится триплет нуклеотидов, соответствующий по коду определенной аминокислоте (антикодон), а основание («черешок») служит местом присоединения этой аминокислоты. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3"-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ) к участку ССА (рис. 5)
Транспортная РНК доставляет аминокислоты к рибосомам. По принципу комплементарности антикодон связывается со своим кодоном, причем аминокислота располагается у активного центра рибосомы и с помощью ферментов соединяется с ранее поступившими аминокислотами. Затем тРНК освобождается от аминокислоты, а молекула иРНК продвигается вперед на один триплет, и процесс повторяется.

Рис. 7.5. Строение молекулы Т-РНК
Различают три этапа в биосинтезе белка: инициацию , элонгацию и терминацию .
В малой субъединице рибосомы расположен функциональный центр рибосомы (ФЦР) с двумя участками - пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три - в пептидильном и три - в аминоацильном участках.
Инициация . Синтез белка начинается с того момента, когда к 5"-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК.
За счет АТФ происходит передвижение инициаторного комплекса (малая субъединица рибосомы, тРНК с метионином) по НТО до метионинового кодона АУГ. Этот процесс называется сканированием .
Элонгация . Как только в Р-участок сканирующего комплекса попадает кодон АУГ, происходит присоединение большой субъединицы рибосомы. В А-участок ФЦР поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислоты.
Скорость передвижения рибосомы по иРНК - 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут.
Терминация . Когда в А-участок попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения, полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.

Рис. 7.6. Процесс трансляции (шаг 1)

Рис. 7.7. Процесс трансляции (шаг 2)

Рис. 7.8. Процесс трансляции (шаг 3)

Рис. 7.9. Процесс трансляции (шаг 4)

Рис. 7.10. Биосинтез белка (общая схема)
Так постепенно наращивается белковая цепочка, в которой аминокислоты располагаются в строгом соответствии с локализацией кодирующих их триплетов в молекуле иРНК. Синтез полипептидных цепей белков по матрице иРНК называется трансляцией (рис. 10).
В клетках растительных и животных организмов белки непрерывно обновляются. Интенсивность синтеза тех или иных специфических белков определяется активностью соответствующих генов, с которых «считывается» иРНК. Следует отметить, что не все гены функционируют одновременно: активность проявляют лишь те, которые кодируют информацию о структуре белков, необходимых для жизнедеятельности организма в данный момент.
Биосинтез белка и генетический код
Определение 1
Биосинтез белка – ферментативный процесс синтеза белков в клетке. В нём участвуют три структурные элемента клетки – ядро, цитоплазма, рибосомы.
В ядре клетки в молекулах ДНК сохраняется информация о всех белках, которые в ней синтезируются, зашифрованная с помощью четырёхбуквенного кода.
Определение 2
Генетический код – это последовательность расположения нуклеотидов в молекуле ДНК, которая определяет последовательность аминокислот в молекуле белка.
Свойства генетического кода таковы:
Генетический код триплетный, то есть каждой аминокислоте соответствует свой кодовый триплет (кодон ), состоящий из трёх расположенных рядом нуклеотидов.
Пример 1
Аминокислота цистеин кодируется триплетом А-Ц-А, валин – триплетом Ц-А-А.
Код не перекрывается, то есть нуклеотид не может входить в состав двух соседних триплетов.
Код вырожден, то есть одна аминокислота может кодироваться несколькими триплетами.
Пример 2
Аминокислота тирозин кодируется двумя триплетами.
Код не имеет запятых (разделительных знаков), считывание информации происходит тройками нуклеотидов.
Определение 3
Ген – участок молекулы ДНК, который характеризуется определённой последовательностью нуклеотидов и определяет синтез одногой полипептидной цепи.
Код является универсальным, то есть единым для всех живых организмов – от бактерий до человека. У всех организмов есть одни и те же 20 аминокислот, которые кодируются одними и теми же триплетами.
Этапы биосинтеза белка: транскрипция и трансляция
Структура любой белковой молекулы закодирована в ДНК, которая не участвует непосредственно в её синтезе. Она служит лишь матрицей для синтеза РНК.
Процесс биосинтеза белка происходит на рибосомах, которые расположены преимущественно в цитоплазме. Значит, для осуществления передачи к месту синтеза белка генетической информации из ДНК нужен посредник. Эту функцию выполняет иРНК.
Определение 4
Процесс синтеза молекулы иРНК на одной цепи молекулы ДНК на основании принципа комплементарности называется транскрипцией , или переписыванием.
Транскрипция происходит в ядре клетки.
Процесс транскрипции осуществляется одновременно не на всей молекуле ДНК, а лишь на её небольшом участке, который отвечает определённому гену. При этом происходит раскручивание части двойной спирали ДНК и короткий участок одной из цепей оголяется – теперь он будет выполнять роль матрицы для синтеза иРНК.
Потом вдоль этой цепи двигается фермент РНК-полимераза, соединяющий нуклеотиды в цепь иРНК, которая удлиняется.
Замечание 2
Транскрипция может одновременно происходить и на нескольких генах одной хромосомы и на генах разных хромосомах.
Образованная в результате иРНК содержит последовательность нуклеотидов, которая является точной копией последовательности нуклеотидов на матрице.
Замечание 3
Если в молекуле ДНК есть азотистое основание цитозин, то в иРНК – гуанин и наоборот. Комплементарной парой в ДНК является аденин – тимин, а РНК вместо тимина содержит урацил.
На специальных генах синтезируются и два другие типа РНК – тРНК и рРНК.
Начало и окончание синтеза всех типов РНК на матрице ДНК строго фиксированы специальными триплетами, которые контролируют запуск (инициирующие) и остановку (терминальные) синтеза. Они выполняют функции «разделительных знаков» между генами.
Соединение тРНК с аминокислотами происходит в цитоплазме. Молекула тРНК формой напоминает листик клевера, на его верхушке расположен антикодон – триплет нуклеотидов, который кодирует аминокислоту, которую переносит данная тРНК.
Сколько видов аминокислот, столько существует и тРНК.
Замечание 4
Поскольку много аминокислот могут кодироваться несколькими триплетами, то количество тРНК больше 20 (известно около 60 тРНК).
Соединение тРНК с аминокислотами происходит с участием ферментов. Молекулы тРНК транспортируют аминокислоты к рибосомам.
Определение 5
Трансляция – это процесс, в результате которого информация о структуре белка, записанная в иРНК в виде последовательности нуклеотидов, реализуется в виде последовательности аминокислот в молекуле белка, которая синтезируется.
Этот процесс осуществляется в рибосомах.
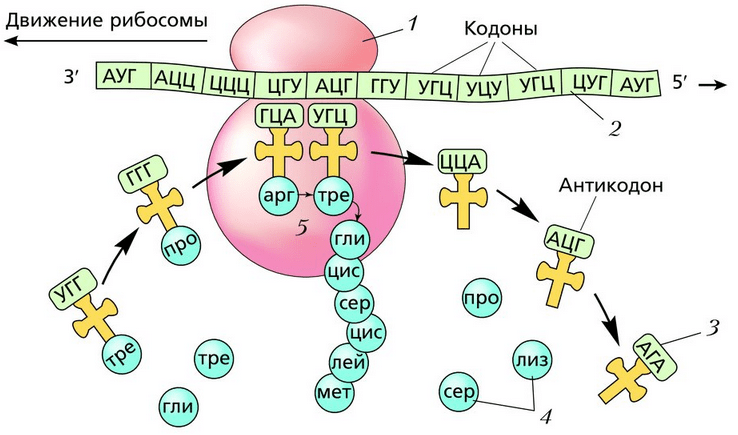
Сначала иРНК присоединяется к рибосоме. На иРНК «нанизывается» первая рибосома, которая синтезирует белок. По мере продвижения рибосомы на конец иРНК, который освободился, «нанизывается» новая рибосома. На одной иРНК могут находиться одновременно более 80 рибосом, которые синтезируют один и тот же белок. Такая группа рибосом, соединённых с одной иРНК, называется полирибосомой , или полисомой . Вид белка, который синтезируется, определяется не рибосомой, а информацией, записанной на иРНК. Одна и та же рибосома способна синтезировать разные белки. После завершения синтеза белка рибосома отделяется от иРНК, а белок поступает в эндоплазматическую сеть.
Каждая рибосома состоит из двух субъединиц – малой и большой. Молекула иРНК присоединяется к малой субъединице. В месте контакта рибосомы и иРН находятся 6 нуклеотидов (2 триплета). К одному из них всё время подходят из цитоплазмы тРНК с разными аминокислотами и касаются антикодоном кодона иРНК. Если триплеты кодона и антикодона оказываются комплементарными, между аминокислотой уже синтезированной части белка и аминокислотой, которая доставляется тРНК, возникает пептидная связь. Соединение аминокислот в молекулу белка осуществляется с участием фермента синтетазы. Молекула тРНК отдаёт аминокислоту и переходит в цитоплазму, а рибосома передвигается на один триплет нуклеотидов. Так последовательно синтезируется полипептидная цепь. Продолжается всё это до тех пор, пока рибосома не дойдёт к одному из трёх терминирующих кодонов: УАА, УАГ или УГА. После этого синтез белка прекращается.
Замечание 5
Таким образом, последовательность кодонов иРНК определяет последовательность включения аминокислот в цепь белка. Синтезированные белки поступают в каналы эндоплазматического ретикулюма. Одна молекула белка в клетке синтезируется за 1 - 2 минуты.